© 2023 John Mattick and Paulo Amaral.
Open Access: This content is Open Access under the Creative Commons license CC-BY-NC-ND.
Bookshelf ID: NBK595927DOI: 10.1201/9781003109242-16

An official website of the United States government

NCBI Bookshelf. A service of the National Library of Medicine, National Institutes of Health.
Mattick J, Amaral P. RNA, the Epicenter of Genetic Information: A new understanding of molecular biology. Abingdon (UK): CRC Press; 2022 Sep 20. doi: 10.1201/9781003109242-16

Expanding from the initial findings that individual RNAs can act as guides, allosteric regulators, scaffolds or decoys, RNA is being shown to have fundamental and highly versatile roles in all aspects of gene regulation in plants and animals, frequently in conjunction with repetitive sequences. RNA regulates chromosome structure through interaction with transposon-derived elements (TEs). Chromatin-remodeling proteins, sometimes referred to as ‘pioneer’ transcription factors, have little or no sequence specificity but bind RNA and address different loci in different cells at different developmental stages. The largest class of sequence-specific transcription factors, containing zinc-finger motifs, also address target loci differentially, bind RNA as well as DNA, and many have higher affinity for RNA-DNA hybrids than for double-stranded DNA. RNA-DNA hybrids and RNA-DNA-DNA triplexes are common in eukaryotic chromatin. Half of the C2H2 zinc-finger proteins in the human genome contain KRAB domains, many primate-specific, which bind TEs. DNA methylation has been known for decades to be RNA-guided. Histone-modifying proteins also have no intrinsic sequence specificity but bind RNA ‘promiscuously’, i.e., bind to many different RNAs. Enhancers express RNAs in the cells in which they are active, and enhancer RNAs are required for enhancer action, which involves chromatin ‘looping’ to form transcriptional hubs. Enhancers have all the signatures of genes, except that they do not encode proteins, and the number of enhancers is approximately the same as the number of lncRNAs expressed from the human genome, which resolves the G-value enigma. Most proteins involved in regulating gene expression in plants and animals, including Hox proteins, transcription factors and histone modifiers, contain ‘intrinsically disordered regions’ (IDRs), the fraction of which increases with developmental complexity. IDRs interact with RNAs to form biomolecular condensates, such as nucleoli and paraspeckles, which are deployed widely to organize specialist subnuclear and cytoplasmic domains, and likely topologically associated domains in chromatin. RNA-IDR interactions may comprise the third dimension of the ancestral protocell. LncRNAs have a modular and highly alternatively spliced structure, with many domains derived from TEs. LncRNAs appear to act as both scaffolds and guides for ribonucleoprotein complexes, a highly efficient and flexible system that, like RNAi and CRISPR, uses RNA signals to direct generic protein effectors to their sites of action, to program development and underpin adaptive radiation.
DNA and proteins have been the focus of the study of chromosome structure, but RNA is also a major component, essential to the organization of chromatin and the ‘nuclear matrix’. 1–18 As long ago as 1989, Sheldon Penman and colleagues demonstrated that transcription is required to maintain nuclear structure, and that chromatin integrity is destroyed by treatment with RNase, noting that “ribonucleoprotein granules were dispersed throughout the euchromatic regions” and suggesting “that RNA is a structural component of the nuclear matrix, which in turn may organize the higher order structure of chromatin” 4 (Chapter 4).
Genome-wide mapping and sequencing studies subsequently showed that there are many chromatin-bound RNAs in animal cells and that the locations of long non-coding RNAs in chromatin are “focal, sequence-specific and numerous”, 19 with thousands of “tightly associated” non-coding RNAs tethered adjacent to active genes. 11 , 20 , 21 Well-studied non-coding RNAs such as 7SK, U1, B2 and Alu RNAs, Gas5 and SRA, and more recently a large coterie of enhancer-derived and other lncRNAs, have been shown to be involved in the regulation of transcription initiation, elongation, termination and splicing. 22–25 The stress response induces the transcription downstream of protein-coding genes of thousands of lncRNAs that remain chromatin bound. 26 Chromatin-associated RNAs, which include those transcribed from enhancers and repeats, have been shown to have roles in genome organization via enhancer-promoter interactions and the formation of transcription hubs, heterochromatin and nuclear bodies (or ‘granules’) 11–13 , 15 , 16 , 21 , 27–30 through their interaction with proteins containing intrinsically disordered regions and the formation of phase-separated domains, as set out below.
The scaffolding of euchromatin involves highly abundant (‘CoT1′) repeat RNAs, predominantly from 5′ truncated LINE elements, 31 , 32 the expression of which varies during development and is regulated by other RNAs. 33 , 34 Chromatin-associated RNA proximity ligation reveals an RNA-DNA contact map similar to that observed by DNA-DNA ligation in topologically associated domains. 13 LncRNAs have been shown to regulate TAD formation, 35–37 and a recent analysis identified more than 10,000 RNA–chromatin interactions mediated by protein-coding RNAs and non-coding RNAs. 38 The RNAi machinery has also been shown to regulate nuclear topology. 39 , 40
Many binding sites for CTCF, a zinc-finger containing protein (see below) that appears to anchor boundary sequences in TADs 41 (Chapter 14), are derived from transposable elements 42 and transcriptionally active HERV-H retrotransposons demarcate TADs in human pluripotent stem cells. 43 Similar to that observed with ‘enhancer’ RNAs (see below), lncRNAs have been reported to regulate neighboring genes through interaction with the Mediator complex, 44 , 45 a master coordinator of transcription and cell lineage commitment that also organizes chromosome topology (Chapter 14).
LINE- and centromere-derived repeat RNAs are structural and functional components of centromeric chromatin. 46–49 Heterochromatin formation generally requires the expression of repetitive sequences 50 and the RNAi pathway, 51–55 and RNA binding is required for heterochromatic localization of HP1 and the Suv39h histone methyltransferase. 56–59 Chromatin compaction is also controlled by lncRNAs that target IAP retrotransposons. 60 Telomere formation and maintenance requires specialized non-coding RNAs, 61 , 62 as does pairing of homologous chromosomes in meiosis 63 , 64 and many, if not most, chromatin-associated proteins bind RNA, 65 including those involved in other chromatin-regulated process such as DNA stability and damage repair. 66
Chromatin structure is modulated during development by ‘pioneer transcription factors’ that alter cell fate in plants and animals by targeting nucleosomes and/or common DNA motifs. 67–71 The best known examples of reprogramming proteins are the ‘Yamanaka’ factors, Oct4 (Pou5f1 gene), Sox2, Klf4 and c-Myc, which are (collectively) capable of converting differentiated cells to ‘induced pluripotent stem cells’ (iPSCs), 72–74 a process enhanced by inclusion of the RNA-binding protein, Lin28. 75 Oct4 is also involved, inter alia, in the differentiation of pluripotent cells to form the cranial neural crest. 76
Another key pluripotency and reprogramming factor is Nanog, a homeobox-containing protein. 77 , 78 Homeoboxes are helix-loop-helix DNA-binding domains that exhibit a preference, but not specificity, for the common motif TAAT, 79 , 80 in the case of Nanog TAAT(G/T)(G/T). 81 Oct4 is also a homeobox-containing protein that recognizes the loose consensus sequence TTT(G/T)(G/C)(T/A)T(T/A), which occurs at thousands of sites around the genome. 82–84
The expression of Oct4, Nanog and other pluripotency factors a is regulated by non-coding RNAs, b including pseudogene-derived lncRNAs, 93–98 one of which recruits the histone-lysine N-methyltransferase SUV39H1 to epigenetically silence Oct4 expression. 97 , 98 Reciprocally Oct4 and Nanog regulate the expression of lncRNAs that modulate pluripotency. 99 Oct4 and Nanog also have multiple pseudogenes, 100–103 some of which are differentially expressed in pluripotent and tumor cell lines. 102 , 104
There are 16 classes of genes encoding homeobox proteins in animals, 11 in plants, with hundreds of orthologs in the human genome, most of which contain additional domains. 80 As noted already (Chapter 5), Hox proteins are ‘master controllers’ of gene expression patterns during animal and plant development, and regulate the expression of many genes at different developmental stages. While they recognize similar sequences in vitro, Hox proteins display wide functional diversity and identification of their in vivo genomic targets has proven elusive, as has the identification of the targets of Oct4 and Sox2. 80 , 83 , 105–109 Analysis of genome-wide DNase I hypersensitivity profiles and transcription factor (TF)-binding sites identified 120 and validated eight ‘pioneer’ TF families that dynamically open chromatin (including Sox2, Oct4 and Hoxa11), and identified ‘settler’ TFs (including c-Myc), and the nuclear hormone receptor RXR:RAR and NF-κB families, whose genomic binding is dependent on chromatin opening by pioneer TFs. 110
The targets of Sox2, Oct4, Nanog and other Hox proteins change with developmental stage, 77 , 111 , 112 all of which suggests that other factors are involved in determining their locus specificity. In this context, it may not be an outlier observation that the Drosophila Hox protein Bicoid (which controls anterior-posterior patterning) binds RNA via its homeodomain, 113 , 114 nor that highly conserved lncRNAs are produced in vertebrate endoderm lineages from paralogous regions in HOXA and HOXB clusters. 115
Sox2 is a member of a subclass of ‘high mobility group’ (HMG) proteins, the most abundant chromatin-associated proteins after histones. HMG proteins bend DNA structure, initiate chromatin opening and facilitate nucleosome remodeling. 116–118 There are three classes, one of which (HMG-A) is abundant in embryonic cells and binds AT-rich sequences, another (HMG-N) binds nucleosomes, and the third (HMG-B, which includes the Sox proteins) binds the DNA helix minor groove with no sequence specificity. 119 , 120 Sox2 influences development not only in pluripotent stem cells but also in the lung, ear and eye, and in neural lineages, but how it and other HMG-B proteins achieve their tissue-specific versatility is unclear. 107
While Sox2 has low affinity for DNA, 116 it binds RNA with high affinity through its HMG domain, 121 , 122 as do other members of the HMG-B family, 123 “which requires a reassessment of how these proteins establish proper patterns of gene expression across the genome”. 121 The HMG-B domain of the mammalian sex-determining protein Sry is homologous to the RNA-binding domain of a viral protein, 124 suggesting that their target selection in vivo is guided by trans-acting RNA signals. There are well-documented examples of lncRNAs that interact with Sox2 to regulate pluripotency, neurogenesis, neuronal differentiation and brain development, 122 , 125–128 and a lncRNA has been shown to interact with a chromatin-remodeling complex to induce nucleosome repositioning. 129
Sox2, Nanog and Oct4 are often found at super enhancers 130 and state-specific differences in enhancer activity correspond with reconfiguration of Sox2, Nanog and Oct4 binding and target gene expression. 111 The lncRNA Evf2 selectively represses genes across megabase distances by coupling recruitment and sequestration of Sox2 into phase-separated domains (see below), affecting enhancer targeting and activity, with genome-wide effects. 122 In human embryonic stem cells Oct4 and Nanog associate with transcripts of the human endogenous retrovirus subfamily H (HERV-H) transposable elements, which are required to maintain stem cell identity and whose terminal repeats function as enhancers. 131 , 132
The classic master switch transcription factor, MyoD, which can reprogram fibroblasts into muscle cells and is central to muscle differentiation in vivo, 133 is regulated by lncRNAs, 134–136 as are other aspects of muscle gene expression. 137–139 The pioneer transcription factor CBP also binds RNAs, including those transcribed from enhancers, to stimulate histone acetylation and transcription. 140
Nucleosome repositioning and remodeling is accomplished by the ATP-dependent imitation switch (ISWI), chromodomain helicase DNA-binding (CHD), SWI/SNF (switch/‘sucrose non-fermentable’) (SWI/SNF) and INO80 complexes. 141 , 142 These complexes are directed to specific sites in chromatin or antagonized by lncRNAs, including Xist and enhancer RNAs, in processes as diverse as rRNA synthesis, myogenic differentiation and proliferation, endothelial proliferation, migration and angiogenic function, atherosclerosis, cardiomyopathy, liver regeneration and stem cell renewal, immunity and inflammation, and various cancers, 129 , 136 , 143–160 leading one group to conclude that “every cell type expresses precise lncRNA signatures to control lineage-specific regulatory programs”. 160
However, the patchy data on the binding of RNAs by the various proteins that control chromatin remodeling during development reflects limited investigations because of the expectation that all ‘transcription factors’ bind to DNA, rather than be directed by RNA-DNA and other RNA-mediated interactions.
Loose DNA sequence specificities are a feature of eukaryotic transcription factors generally. While eukaryotic genomes are orders of magnitude larger than those of prokaryotes, their more conventional TFs have shorter DNA recognition sites (6–10bp versus 15–25bp in E. coli 161 ), often expressed as a ‘consensus’ sequence, but better represented by multiple sequences, with many TFs recognizing different primary and secondary motifs. 162–166
Moreover, high-throughput chromatin immunoprecipitation experiments with antibodies against specific TFs show different patterns of binding in different cell types, so additional factors must be involved. Such factors can be either (or both) trans-acting signals or chromatin accessibility, the latter supported by the observation that TF-binding sites are nucleosome depleted and DNase-sensitive, indicating that epigenomic decisions precede TF factor binding. 167–170
The largest class of TFs in animals and plants contain ‘zinc-finger’ (ZF) domains, c specifically the C2H2 class, of which there are over 700 encoded in the human genome, and which recognize more sequence motifs than all other transcription factors combined. 171 Human C2H2-ZF proteins contain an average ~10 C2H2 domains (ranging from 1 to 30), classified into three groups: ‘triple’, ‘multiple-adjacent’, and ‘separated-paired’ C2H2 finger proteins, enabling some to bind multiple ligands.
It is thought that most ZFs bind to DNA, although most of the binding sequences are unidentified, 165 but many ZFs also bind to RNA or protein, and some to RNA only. 162 , 172 , 173 The classic example is TFIIIA (Chapter 8), which is required for the transcription of 5S rRNA genes and is titrated off DNA by its higher affinity for 5S rRNA, the first demonstration of the regulation of TFs by RNAs. 174 , 175
A large fraction of C2H2-ZF TFs have been shown to regulate alternative splicing. 176 A splice variant that introduces three additional amino acids (KTS) between the third and fourth ZFs of the Wilm’s tumor protein WT1 d changes the specificity of the WT1 protein from DNA to spliceosomes, 178 presumably by binding RNA, given that WT1 also contains an RNA recognition motif 179 and transcription and splicing are coupled. 180 Disturbance of the ratio of +/-KTS isoforms causes a developmental syndrome, affecting kidney and genital development. 181 Both isoforms bind DNA and RNA in vitro, 182–184 shuffle between the nucleus and translating polysomes in the cytoplasm, 185 and their subnuclear location is RNase- but not DNase-sensitive. 182
A 1994 analysis by Yigong Shi and Jeremy Berg of two representative C2H2-ZF proteins, one of which was Sp1 (which controls the expression of many housekeeping, tissue-specific, cell cycle and signaling pathway response genes 186 ), showed that they have a higher affinity for RNA-DNA hybrids than for double-stranded DNA and that this increased affinity was strand-specific, i.e., dependent on which strand is RNA. 187
The C2H2-ZF transcription factor YY1, which regulates the expression of various genes during embryogenesis, cell differentiation and proliferation, 188 binds chromatin in an RNA-enhanced fashion 189 and appears to play a major role in mediating enhancer-promoter loops. 190 YY1 also interacts with an RNA-binding protein involved in splicing regulation, depletion of which attenuates YY1 chromatin binding and YY1-dependent DNA looping and transcription. 29 The ZF-containing TAD insulator CTCF has also been shown to be a high-affinity RNA-binding protein. 65 , 191–195
Later studies confirmed that over 800 human proteins bind RNA-DNA hybrids and over 300 prefer binding RNA-DNA hybrids over dsDNA. 196 These observations raise the possibility, if not the likelihood, that trans-acting RNAs are involved in the exposure and selection of genomic TF-binding sites, explaining the differential locus specificity of TF binding and the reason for a loose consensus sequence, e as well as enabling directionality of action by strand selection.
RNA-DNA hybrids f (which form ‘R-loops’ with the displaced DNA strand) occur widely throughout the human genome 198 , 199 and even encompass 8% of the yeast genome. 200 RNA-DNA hybrids are enriched at unmethylated CpG-rich promoters, transcription start sites and regions enriched for activating histone modifications such as H3K4me1/2/3, H3K9ac and H3K27ac. 201 RNA-DNA hybrids regulate genome stability and DNA repair, 197 , 202 , 203 promoter-proximal chromatin architecture and cellular differentiation, 204 transcriptional activation 205 and “are enriched at loci with … potential transcriptional regulatory properties … supporting a model of certain transcription factors binding preferentially to the RNA:DNA conformation”. 206 The formation and stability of RNA-DNA hybrids are in turn regulated by RNA methylation and other modifications 197 , 207 (Chapter 17).
Nucleic acid triplex structures g (wherein a single stranded DNA or RNA forms ‘Hoogsteen’ hydrogen bonds with the purine-rich strand of polypyrimidine-polypurine tracts in the major groove of duplex DNA) also occur in vivo, as first detected by the sequence-specific binding of RNA to ‘native’ dsDNA. 214 Triplex-forming sequences are overrepresented in eukaryotic but not bacterial genomes, notably in regulatory regions and promoters. 215–217 Antibody and sequencing studies have also shown that triplex structures abound in eukaryotic chromosomes 218–221 (Figure 16.1). Triplex hotspots targeted by lncRNAs have been proposed to contribute to chromatin compartmentalization in conjunction with ‘architectural’ TFs such as CTCF 222 and the positions of lncRNA:DNA triplex-forming sites have been shown to be predictors for TADs. 223 Triplex-forming oligonucleotides have been shown to alter cell division, inhibit tumor growth, stimulate recombination and modulate target gene expression. 224–227
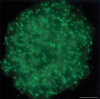
Triplex-forming DNAs in the interphase nucleus of a human monocytic leukemia cell visualized in situ by an anti-triplex monoclonal antibody. The bar represents 5 μm. (Reproduced from Ohno et al. with permission of Springer Nature.)
Many lncRNAs, including those expressed from enhancers, have been shown to interact sequence-specifically with DNA 228 to regulate various processes through R-loop or triplex formation, including chromatin architecture, transcription, radiation response, cell proliferation, cell differentiation and organ development, in some cases (at least) intersecting with epigenetic pathways. 204 , 217 , 229–240 Triplexes are also involved in small RNA-mediated transcriptional gene silencing. 241
A good example, from plants, is the lncRNA APOLO, which coordinates the expression of multiple genes in response to cold through sequence complementarity and R-loop formation, decoys Polycomb and binds transcription factors at the promoter of a master regulator of root hair formation. 242–244 Amazingly, APOLO function can be partly mimicked by the sequence-unrelated lncRNA UPAT, which interacts with orthologous proteins in mammals, indicating conservation of regulatory structures and lncRNA functions across kingdoms. 244
The enhancer lncRNA KHPS1 forms a triplex with enhancer DNA sequences to activate expression of the neighboring SPHK1 gene, by evicting CTCF, which insulates the enhancer from the SPHK1 promoter. Deletion of the triplex-forming sequence attenuates SPHK1 expression, leading to decreased cell migration and invasion, and the targeting of KHPS1 lncRNA can be switched by swapping the triplex-forming promoter sequence to other genes. 232 , 233
Other classes of ‘transcription factors’ such as Y-box proteins also bind RNA, and known RNA-binding proteins such as hnRNP K (better known as a ‘splicing factor’) also act as transcription factors. 245 , 246 The ‘paired-box’ transcription factor Pax5, which is a ‘master regulator’ of B-cell development by recruiting chromatin-remodeling, histone-modifying and basal transcription factor complexes to its target genes, 247 is hijacked to the Epstein Barr Virus genome by a viral-encoded non-coding RNA. 248 Another ‘transcription factor’, the nuclear hormone receptor ESR1 (estrogen receptor α), which is commonly activated in breast cancer, is also an RNA-binding protein. 249
Dual RNA-DNA or ambiguous RNA/DNA-binding proteins also include p53, 172 , 250 the ‘guardian of the genome’, possibly the most intensively studied gene and protein in human molecular biology, which binds a lncRNA (‘damage-induced noncoding RNA’, DINO). 251 The dual DNA/RNA-binding protein TLS/FUS (Translocated in LipoSarcoma/FUsed in Sarcoma) is allosterically regulated by lncRNA pncRNA-D. 252–254 Even RNA polymerase is regulated by RNAs. In mammals, RNA polymerase II is repressed by short RNA polymerase III transcripts derived from mouse B2 and human Alu repeat (SINE) elements. 255–258 These elements also provide mobile RNA polymerase II promoters. 259
Importantly, approximately half (~350) of the human C2H2-ZF proteins, many of which are unique to primates, contain a KRAB transcriptional repression domain, which binds TEs, h and evolved by recurrent TE capture that partners them with emergent TE-mediated regulatory networks, influencing genomic imprinting, placental growth and brain development. 171 , 260–270 Most eukaryotic TFs also contain intrinsically disordered domains that overlap their DNA-binding domain and direct their target specificity, 271–275 likely by interaction with guide RNAs (see below).
Presumably, different types of DNA/RNA-binding proteins recognize different types of nucleic acid structures and transact a different type of signal in different contexts within the decisional systems that control cell division and differentiation during development, as well as in physiological responses. The fact that most eukaryotic ‘transcription factors’ have confusing and enigmatic functions attests to the likelihood that they have been interpreted in the wrong conceptual framework, with RNA the missing link. 276
Transcriptional gene silencing in fungi and plants by RNA-directed DNA methylation was well established in the 1980s and 1990s (Chapter 12). These studies eventually showed that the enzymes that methylate DNA are directed to their sites of action by small RNAs interacting with the RNAi protein AGO4. 277 , 278 Small RNAs (miRNAs, siRNAs and piRNAs) also induce site-specific DNA methylation in animals, 52 , 53 , 279 , 280 which again involves Argonaute proteins, 281–285 suggesting that what had originated as an RNA-based mechanism for defense against viruses has been co-opted as a means of genome regulation. i
In 2004, Linda Jeffery and Sara Nakielny showed that the de novo DNA methylases Dnmt3a and Dnmt3b, but not the maintenance methylase Dnmt1, bind siRNAs with high affinity. 287 Later others reported that Dnmt1 (which restores methylation at hemi-methylated CpG sites after DNA replication) binds lncRNAs to alter DNA methylation patters at cognate loci. 288–291
Demethylation also appears to be an active process guided by RNAs. 292–295 RNA-directed DNA demethylation has also been reported to involve R-loop formation, 239 , 294 and recruitment of the TET2 dioxygenase/demethylase (which unlike other TET enzymes does not contain a DNA-binding domain, but does bind RNA 296 , 297 ) by RNAs transcribed from endogenous retroviruses. 298
Some methyl-CpG-binding proteins, including MeCP2, bind siRNAs and other RNAs, mediated through a domain distinct from the methyl-CpG-binding domain with, interestingly, RNA and methyl-CpG binding being mutually exclusive, 287 , 299 although there seems to be variations of the regulatory mechanisms, including RNA-mediated recruitment to phase-separated heterochromatin compartments (see below). LncRNAs have also been shown to link DNA methylation with histone modification through triplex formation with target sequences. 300
A range of histone variants and over 100 different histone modifications are differentially incorporated into nucleosomes located at millions of different positions in different cell types and different stages of development and differentiation (Chapter 14). However, like DNA methylation enzymes, histone-modifying enzymes also have no intrinsic DNA-binding capacity or specificity, which is often assumed to be provided by sequence-specific DNA-binding proteins or transcription factors that interact with them. On the other hand, like DNA methylation enzymes, many histone modification writers and readers contain domains that bind RNA and/or contain RNA-binding modules. These include RNA recognition motifs, 301 chromodomains, 296 , 302 , 303 bromodomains, 296 Tudor domains, 304 PRC2 subunits EZH2, EED, Suz12 and Jarid2, 305–310 the H3K20 trimethylase Suv4–20h 60 and other histone-modifying complexess. 311
RNA binding to histones was first reported in the mid-1960s 312 , 313 (Chapter 4). Around the turn of the century, a number of groups showed that PcG (Polycomb group) proteins from C. elegans and vertebrates also bind RNA, and that this binding is essential for their chromatin localization and repression of homeotic genes. 314–316 In 2005, Renato Paro and colleagues showed that the switch from the silenced to the activated state of a Polycomb response element in the Drosophila bithorax locus (Chapter 5) requires non-coding transcription. 317
In 2007, John Rinn and colleagues showed that lncRNAs transcribed from human homeotic gene loci, like those in Drosophila, are expressed along developmental axes and demarcate active and silent chromosomal domains that have different H3K27me3 profiles and RNA polymerase accessibility, the exemplar of which, HOTAIR (Chapter 13), interacts with PRC2 and is required for PRC2 occupancy and histone H3K27 trimethylation at the HOXD locus. 318 Other studies showed, for example, that retinoic acid-induced expression of lncRNAs follows the collinear activation state and correlates with loss of Polycomb repression at the HOXA locus. 319
In 2008, the groups of Chandrasekhar Kanduri and Peter Fraser showed that lncRNAs differentially expressed from parentally imprinted loci, specifically the 105 kb Air RNA and the 91 kb Kcnq1ot1 RNA (formerly KvLQT1-AS, Chapter 9), and later others, also bind PRC2 to repress the relevant alleles. 320–322 In the same year, we showed that lncRNAs expressed from the antisense strand of homeotic gene loci during embryonic stem cell differentiation are associated with both the chromatin-activating Trithorax MLL1 complex and activated chromatin containing H3K4me3 marks. 323 Subsequently other lncRNAs (including enhancer RNAs and small RNAs derived from them) were also shown to associate with Trithorax complexes, including RNAs involved in maintenance of stem cell fates and lineage specification (such as Evx1-as and HOTTIP), 14 , 45 , 324–334 and even grain yield in rice. 335
In 2009, Rinn and colleagues surveyed over 3,300 lncRNAs and showed that ~20% (but only ~2% of mRNAs) interact with PRC2, and that others are bound by other chromatin-modifying complexes. 336 Moreover, knocking down a selection of these RNAs caused derepression of genes normally silenced by PRC2. 336 Over 9,000 RNAs bind PRC2 in embryonic stem cells, 306 with many individual cases, including the lncRNAs H19, MEG3, ANRIL j and HOTAIR, subsequently characterized in some detail. 303 , 309 , 337–342
RNA has also been shown to be required for PRC2 chromatin occupancy, PRC2 function and cell state definition. 343 Short RNAs transcribed from Polycomb-repressed loci resemble PRC2-binding sites in Xist, and interact with PRC2 through its subunit Suz12. 307 PRC2 binds G-quadruplex structures in RNA, 344 which inhibit PRC2 activity and are antagonized by allosteric activation of PRC2 by H3K27me3 and regulators of histone methyltransferases, 345 , 346 indicating complex decisional transactions.
PRC1 function also appears to be controlled by RNA 303 and PRC1 resides in membrane-less phase-separated nuclear organelles 347 that are likely to be RNA nucleated (see below).
PRC2 k binds many RNAs ‘promiscuously’, 348 a description l that does not mean ‘non-specifically’. 339 , 349 The association of Polycomb and Trithorax complexes with many RNAs, likely through orthologous domains (see below), is consistent with their function as guide molecules for RNA-directed site-specific histone and DNA modifications. It is also consistent with the fact that Polycomb and Trithorax proteins are involved in many differentiation and developmental decisions, from cell cycle regulation to embryogenesis and body plan specification, 350–355 so their binding to many different guide RNAs would be expected. Again, these include RNA-DNA, RNA-RNA and RNA-protein interactions, recruitment or eviction of histone modifiers and other chromatin-modifying proteins, alteration of DNA topology, allosteric inhibition, and reorganization of phase-separated domains. 342 , 343 , 345 , 346 , 356–364
LncRNAs also control switching between Polycomb and Trithorax response elements. 329 , 356 Other histone modifications are also regulated by lncRNAs, m including during memory formation. 366 An intriguing observation, not inconsistent with RNA involvement, is that histone-modifying enzymes, rather than the parental histones, may remain associated with DNA through replication to re-establish the epigenetic information on the newly assembled chromatin. 367
While initially thought to be a special case, the best characterized and most illustrative example of the complex interplay between lncRNAs, chromatin structure and gene expression is Xist. 368 , 369 Xist has eight exons and is 17 kb in length. 370 It has a highly modular structure, including a number of types of conserved ‘repeat’ sequences, and interacts with over 80 different proteins including cohesin, Polycomb and other chromatin remodelers, 370–374 at low copy number. 375 , 376
Xist-mediated silencing of the inactive X chromosome in mammals requires its repeat sequences 377–382 and involves Polycomb recruitment, deacetylation of H3K27ac and H3K27 methylation on the silenced chromosome. 305 , 368 , 382–387 Spreading involves the partitioning of chromatin topology 388–392 by the formation of phase-separated domains 376 , 393 , 394 and the interaction with RNA-binding proteins with repetitive elements (in particular LINE-1 elements) in the X chromosome to recruit silencing mechanisms targeted to repeats, 379 , 394–399 as first proposed by Mary Lyon, 400 , 401 likely via triplex formation. 402 Xist also acts as a suppressor of hematological cancers 403 and is essential to maintain X-inactivation of immune genes, dysregulated in females suffering systemic lupus erythematosus or COVID-19 infection. 385
Xist expression and action is controlled and effected by other lncRNAs 369 that are expressed antisense to Xist (Tsix, which blocks RepA RNA binding to PRC2 305 , 404 , 405 ) or from adjacent loci on the active or inactive X chromosomes 406–408 (Figure 16.2). The Jpx lncRNA, whose gene resides ∼10 kb upstream of Xist, activates Xist by regulating CTCF anchor site selection to alter the topography of chromosome loops. 37 , 409 , 410 A primate-specific TE-derived lncRNA, XACT, coats active X chromosomes in pluripotent cells and is connected into the pluripotency regulatory network in humans by primate-specific retroviral enhancer. 411 Another lncRNA expressed from the X-inactivation center, Tsx, functions in germ and stem cell development as well as in learning and behavior. 412 The lncRNA Firre, which also contains a number of repeats, one of which interacts with the nuclear matrix factor hnRNPU, anchors the inactive X chromosome near the nucleolus and is required for the maintenance of its repressive H3K27me3 marks. 413 Firre is also required for the topological organization of other chromosomal regions, 414 , 415 and is involved in other developmental processes including adipogenesis and hematopoiesis. 416
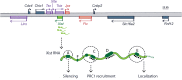
The organization of the Xist locus. The Xist, Tsix, Jpx, Xite, Tsx and Ftx genes specify lncRNAs. (Reproduced from Loda and Heard under Creative Commons attribution license.)
X-inactivation also involves the RNAi enzyme Dicer 368 , 417 , 418 and methylation of Xist transcripts, 419 implicating small RNAs, the RNA interference pathway and RNA modifications in a complex set of decisional pathways that control chromatin architecture from yeast to humans. 420 , 421 Chromosomal dosage compensation in Drosophila, which involves global activation of the single X chromosome in males is controlled by the lncRNAs roX1 and roX2, 422 via a conserved predicted stem-loop structure required for histone H4K16 acetylation of the X chromosome 423 and selective X-chromosome subnuclear compartmentalization. 424
As discussed in Chapter 14, enhancers play a key role in specifying cell identity and are a signature feature of the regulation of gene expression during development.
Enhancers were initially identified by their activity, rather than their physical manifestation, but have been interpreted in terms of the initial speculations about the latter, postulated and promulgated by Mark Ptashne (and widely accepted) to comprise cluster of binding sites for TFs that act at a distance by ‘looping’ to make contact with the promoters of target genes, some of which can be located hundreds of kilobases distant. 425–431
However, early studies had shown that lncRNAs are transcribed from enhancer regions in well-studied loci, 432–434 with supporting evidence accumulating with improving transcriptomic and chromatin analysis technologies. Although sometimes referred to as (protein-coding) ‘gene deserts’, 435 enhancers exhibit the characteristics of bona fide genes, including nucleosome-depleted promoter regions that bind transcription factors and the transcription of adjacent sequences. 436–441 Indeed, the epigenetic architecture of, and the features of transcription initiation at, the promoters of conventional protein-coding genes and enhancers are almost indistinguishable. 436 , 441–443
Enhancers and ‘super-enhancers’ are transcribed to produce non-coding RNAs specifically in the cells in which they are active 317 , 435 , 437 , 441–447 and their expression is considered the best molecular indicator of enhancer activity in developmental processes 437 , 443 , 448–453 and cancers. 91 , 454–457
Enhancers recruit RNA polymerase 458 and produce short unstable bidirectional transcripts (‘eRNAs’) from their promoters, 437 , 444 , 459–462 as do protein-coding genes, 430 , 463–465 and it is uncertain whether these transcripts play a role in enhancer action or simply mark active promoters and/or reflect promiscuous RNA polymerase initiation at accessible chromatin. 441 , 461 , 464 , 466–468 On the other hand, enhancers also express multi-exonic lncRNAs, the half-life of which is exosome regulated, 466 and many if not most lncRNAs likely derive from enhancers. 441 , 444 , 446 , 469–479 Enhancers with tissue-specific activity are enriched in introns, suggesting that “the genomic location of active enhancers is key for the tissue-specific control of gene expression”. 480
There is good evidence that enhancers regulate chromosome looping and local chromatin reorganization to alter cell fate, 435 , 441 , 451 , 481 , 482 which is consistent with but does not demonstrate direct contact between enhancer TF-binding sites and the promoters of target genes. It is also consistent with enhancer RNAs organizing looping with target genes, which has been demonstrated in at least two cases, 482 , 483 and/or the formation of topologically associated, possibly phase-separated, chromatin domains 16 as local hubs of transcription regulation (Figure 16.3). Recent studies suggest that there is no direct contact between TFs bound at the enhancer and the promoter of genes regulated by enhancer action, 484 and that maintenance of enhancer-promoter interactions and activation of transcription are separable events. 485

Cloud formed by the enhancer lncRNA Evf2 and its localization to activated (Umad1, 1.6 Mb distant) and repressed (Akr1b8, 27 Mb distant) target protein-coding genes. (Reproduced with permission from Cajigas et al with permission of Elsevier.)
At the heart of the debates about the mechanism of enhancer action has been the question of whether the RNAs transcribed from active enhancers are simply a passive by-product of TF occupancy, or whether it is the ‘act of enhancer transcription’ or the enhancer RNAs themselves that mediate enhancer action. 442 , 447 , 486 , 487 The evidence for these possibilities, which are not mutually exclusive, has been widely canvassed and variously interpreted, with the former initially favored because it fitted the TF paradigm of protein-coding gene regulation and did not require acceptance of large numbers of regulatory RNAs. 441 , 476 , 487–489 In line with these preconceptions, some studies have reported that the transcribed enhancer RNA sequences can be partly/substantially (it is difficult to be sure n ) deleted, truncated or replaced with no obvious effect (see, e.g., 490 , 491 ).
Other studies showed that enhancer RNAs are required for enhancer activity. 92 , 473 , 479 , 482 , 492–498 For example, deletion in mice of the multi-exonic lncRNA Maenli, which is expressed from an enhancer that controls limb development and is deleted in a human developmental disorder, recapitulates the human phenotype. 479 Deletion of internal exons of the enhancer-derived lncRNA ThymoD blocks T-cell development and causes developmental malignancies. 482 Truncation of the lncRNA Evf2, which is transcribed from the highly conserved Dlx5/6 enhancer that spatially organizes the expression of a 27 Mb region on chr6 during mouse forebrain development, abrogates the action of the enhancer. 473 , 499 The latter study also showed that the 5′ and 3′ ends of the Evf2 enhancer RNA had different functions, o and that Evf2 formed an “RNA cloud” encompassing its target genes. 473 It has also been shown that methylation and splicing of enhancer RNAs are required for enhancer function and chromatin organization. 490 , 500–502
siRNA-mediated knockdown of enhancer RNA also abrogates or reduces enhancer action, demonstrating the involvement of the RNA. 477 , 503–507 RNA has been shown to be required for the formation of enhancer-target promoter contacts by the transcription factor YY1 190 (which itself regulates the expression of many lncRNAs 508 ) and ectopic expression of enhancer RNAs upregulates expression of the genes normally targeted by the enhancer. 472 , 509
Careful analysis shows that the variable phenotypic consequences of enhancer lncRNA knockdown, truncation or ablation depend on the details. 496 , 510 , 511 For example, a short deletion of the promoter and first two exons of the 17 kb lncRNA Hand2os1, which is expressed from an enhancer essential for heart morphogenesis, did not produce discernable heart phenotypes, but deletion of exons 4 and 5 caused severe contraction defects in adult heart that worsened with age, and deletion of the entire Hand2os1 sequence led to dysregulated cardiac gene expression, septum lesion, heart hypoplasia and perinatal death. 510
It has been shown that enhancer RNAs produced in response to immune signaling bind the bromodomains of BRD4 (and other epigenetic reader bromodomain-containing proteins) to augment BRD4 enhancer recruitment and transcriptional cofactor activity. 512 BRD4 also cooperates with oncogenic fusions of MLL1 to induce transcriptional activation of enhancer RNAs, one of which has been shown to bind histone H4K31ac to promote histone recognition and oncogene transcription. 513 Other mechanisms may involve the interplay between different types of regulatory RNAs and chromatin-associated proteins, as suggested by interactions between NEAT1 and BRD4/WDR5 p complexes, and enhancer RNAs with cohesin, with effects on specific target genes. 474 Finally, there is also evidence of concerted action of cis-acting enhancer RNAs with other transcripts that have trans-acting roles. 514–516
While there may not yet be universal acceptance, the evidence is accumulating that enhancer RNAs are integral to enhancer function, 517 and that enhancer RNAs are simply a (large) class of lncRNAs that regulate chromatin architecture and the expression of protein-coding and (other) lncRNAs, albeit through physical mechanisms that are not yet well understood, but involve recognition of effector proteins and sequence-specific RNA-DNA contacts via R-loops or triplexes, 17 , 221 , 236 , 518 and formation of topologically associated domains to form developmental stage-specific transcriptional hubs. 431 , 519 , 520 It is also evident that transcription itself modulates chromosome topology and phase transition-driven nuclear body assembly. 521–524
There are ~400,000 enhancers (and ∼400,000 differentially accessible chromatin elements, 525 , 526 which likely correspond to promoters) in the human genome. 438 , 441 , 444 , 448 , 456 , 460 , 527–533 This is similar to the number of lncRNAs expressed from the human genome (Chapter 13). Indeed, apart from the fact that they do not encode proteins, q enhancers might be properly viewed as genes, which, together with the multitude of other genes expressing functional non-coding RNAs, resolves the G-value enigma.
mRNAs may also have enhancer function, 12 , 490 which would not be surprising given the interwoven nature of the expression of genetic information during the complex ontogenies that underpin animal and, to a lesser extent, plant development.
It has been known for many years that there are many ribonucleoprotein (RNP) complexes that exist in defined territories in the nucleus and cytoplasm of eukaryotic cells, prominent examples of which include nucleoli, spliceosomes, paraspeckles and stress granules, 537–539 none of which are membrane-bound. One of the most important advances of recent years, first canvassed by Harry Walter and Donald Brooks in 1995, and later demonstrated by Clifford Brangwynne, Anthony Hyman and colleagues, is that these and other focal or ‘punctate’ organelles are phase-separated condensates or ‘coacervates’ 540 , 541 that compartmentalize biochemical and regulatory hubs, 542 although not without some controversy and uncertainty. 543
Phase-separated condensates, which are heterogeneous in constitution and properties, are commonly referred to as ‘phase-separated domains’ (PSDs). They are also called ‘liquid crystal domains’, ‘liquid droplets’, ‘biomolecular condensates’, ‘nuclear clouds’ or ‘nuclear bodies’, 544 and exist in an aqueous state distinct from the surrounding environment, the biological manifestation of liquid or soft matter physics. 545
In vitro PSDs form spontaneously by association of oppositely charged molecules such as negatively charged RNA interacting with positively charged proteins. 544 In vivo they are formed by interactions between RNAs, r RNA-binding proteins and proteins with intrinsically disordered domains (IDRs), 27 , 539 , 542 , 544 , 546–553 explaining the latter’s previously mysterious function. 539
IDRs lack rigid tertiary structure and are characterized by a high proportion of small, polar and positively charged amino acids (arginine, histidine and lysine), often in the form of RGG/RG, histidine-rich domains or other repeats. 554–557 IDRs are promiscuous, i.e., they interact with and are tunable by many partners. 550 , 556–561 Intrinsically disordered RGG/RG domains mediate specificity in RNA binding, 562 , 563 and IDRs flank the DNA-binding domains of transcription factors, 273 direct TF binding and, inter alia, the temporal regulation of transcription complexes that specify neuronal subtypes. 564
IDRs and PSDs occur in bacteria and archaea, 565–568 but there have been sharp increases in the fraction of the proteome containing IDRs between prokaryotes, simple eukaryotes and multicellular organisms, and the number of proteins containing IDRs correlates with the number of cell types, suggesting co-evolution of IDR-mediated transactions with developmental complexity. 559 , 560
IDRs are usually located at the N- or C-terminal region of the protein. IDRs are present in and essential for the function of nearly all of the proteins involved in animal and plant development, s including RNA polymerase, most transcription factors, Hox proteins, histones, histone-modifying proteins, other chromatin-binding proteins, the Mediator complex, RNA-binding proteins, splicing factors, membrane receptors, cytoskeletal proteins and nuclear hormone receptors. 271 , 272 , 274 , 539 , 551 , 560 , 563 , 570–577
Surprisingly, the majority of proteins subject to alternative splicing contain IDRs. 560 Moreover, IDRs are overrepresented in alternatively spliced exons subject to tissue- and lineage-specific regulation, 578–581 especially in exons that are alternatively spliced in mammals but constitutively spliced in other vertebrates, 582 which changes the subcellular localization of the isoform and coordinates phase transitions within the cell. 583 , 584
IDRs also occur in proteins that are flexibly involved in signal transduction and transport, 539 such as the Ras-GTPase-activating proteins (SH3 domain)-binding proteins G3BP (which forms phase-separated domains), 585 clathrin-mediated endocytosis 586 and synapsins, which are required for the maintenance of synaptic vesicle clusters in neurons by IDR-mediated phase separation. 587
IDRs are major sites of post-translational modifications and many biological processes, including the regulation of the cell cycle and circadian clocks, 539 have been shown to be dependent on post-translational modification of IDRs. 539 , 588–590 Post-translational modifications modulate RNA binding 591 and alter the propensity to nucleate PSDs, 592 adding layers of complexity to their interactions and regulation. 539 , 557 , 593 , 594
The known post-translational modifications not only include the 100 or so found in histones, but also 95 in the IDR of the axonal microtubule–associated protein Tau, which is involved in Alzheimer’s and other neurodegenerative diseases. 595 Tandem RNA-binding sites in the RNA-binding protein, TIA-1, facilitate PSD stress granule formation 596 and reduction of this protein protects against Tau-mediated neurodegeneration. 597
Other proteins involved in neurological functions and disorders, such as TDP-43, ataxin, c9orf72 and FMRP (fragile X mental retardation protein), also contain IDRs that are involved in phase separation, controlled in part by post-translational modifications. 598 , 599 For example, the IDR of TDP-43 binds RNAs, 600 and the loss of its RNA-binding ability by mutations or post-translational acetylation leads to its sequestration into PSDs. 601 The IDRs of ataxin mediate formation of neuronal mRNP assemblies, and are essential for long-term memory formation as well as c9orf72-induced neurodegeneration. 602 Neuronal-specific micro-exons overlapping IDRs in the translation initiation factor eIF4G regulate the coalescence of phase-separated granules to repress translation, are misregulated in autism, and their deletion in mice leads to altered hippocampal synaptic plasticity and deficits in social behavior, learning, and memory. 603
Aberrant promiscuity of IDR-containing proteins (IDPs) and perturbations of PSD formation may underlie the dosage sensitivity of oncogenes and other proteins 604 , 605 as well as neurodegenerative disorders such as Alzheimer’s Disease, Parkinson’s Disease, Frontotemporal Dementia, Muscular Dystrophy and Amyotrophic Lateral Sclerosis, where repeat expansions affecting RNA and/or their encoded proteins result in pathological aggregates. 598 , 606 , 607
Mutations in RBPs that cause human monogenic diseases are observed more commonly in IDRs than globular domains, 586 , 608 indicating that, despite their relatively simple composition, IDRs have strong sequence constraints. It is also clear that RNA nucleates the formation, and is the structural scaffold, of PSDs. 27 , 542 , 547 , 549 , 551 , 553 , 609–614
PSDs encompass a range of nuclear compartments: 27 , 615 DNA replication initiation sites; 616 telomeres; 617 centrosomes 618 and meiotic chromosomal pairing foci; 619 , 620 germ granules; 592 , 621 nucleoli, Cajal bodies and ‘histone locus bodies’; 540 , 622–624 ‘extranucleolar droplets’; 523 spliceosomes (‘nuclear speckles’ t ); 626 specialized spliceosomes (via lncRNAs Gomafu u and Malat1); 627–629 paraspeckles v (Neat1); 630–634 heterochromatin; 609 , 635 Polycomb bodies; 347 primate-specific nuclear stress bodies; 636 , 637 nuclear glucocorticoid receptor foci; 638 SARS-Cov2 viral assembly domains; 639 and others, including in plants. 640 They also include cytoplasmic organelles 537 , 641 , 642 such as P-granules, 643 , 644 G-bodies, 645 stress granules, 646 polar bodies (whose formation is dependent on a lncRNA), 647 , 648 localized mRNP translational assemblies 649 , 650 and synaptic compartments. 651
It has been proposed that lncRNAs play a central role in organizing the three-dimensional genome, 521 including the formation of spatial compartments and transcriptional condensates 610 , 614 , 652–655 (Figure 16.4) and hence the four-dimensional patterns of gene expression during differentiation and development. w It has been shown that phase separation drives chromatin looping 657 and is required for the action of enhancers and super-enhancers; 551 , 610 , 658–660 that transcription factors activate genes through the phase-separation capacity of their activation domains by forming PSDs with RNA polymerase II; 661 , 662 that Mediator and RNA polymerase II associate in transcription-dependent condensates; 658 , 661–663 that phase separation of RNA-binding protein promotes polymerase binding and transcription; 664 and that PSDs scaffolded by lncRNAs, including repeat-derived RNAs, mediate heterochromatin formation, 32 , 609 , 614 , 665–669 euchromatin formation, 670 nucleolar structure, 671–674 splicing 675 and DNA damage repair. 676–678

Subcellular and subnuclear localization of RNAs in punctate domains. (Reproduced with permission from Cabili et al. with the permission of the authors under Creative Commons attribution license.)
For example, it has been shown that the cytoplasmic lncRNA NORAD, which is induced by DNA damage and required for genome stability, prevents aberrant mitosis by sequestering Pumilio proteins (which bind many RNAs to regulate stem cell fate, development and neurological functions 680 ) into PSDs via multiple repeats. 681–684 Lack of NORAD accelerates aging in mice. 685 Similarly sequestration of the double-strand beak enzyme RAG1 into nucleoli modulates V(D)J recombination activity. 686 Many natural antisense lncRNAs with embedded mammalian interspersed repeats are overrepresented at loci linked to neurodegeneration and/or encoding IDPs. 687
The PARP1 superfamily, x one of the most abundant proteins in the eukaryotic nucleus, 688–690 which catalyzes the polymerization of ADP-ribose units and attachment of poly (ADP-ribose) polymers to arginines in target proteins, y including histones, for DNA repair, stabilization of replication forks and the modification of chromatin, also binds lncRNAs, 692–695 regulates RNA metabolism 696 , 697 and modulates the phase-separation properties of RNA-binding proteins. 698 , 699
Many lncRNAs appear to be localized to defined nuclear and cytoplasmic foci that resemble liquid droplets, 336 , 414 , 473 , 679 , 700–702 and a genome-wide study identified hundreds of non-coding RNAs forming nuclear compartments near their transcriptional loci, in dozens of cases guiding cooperating proteins into these 3D compartments and regulating the expression of genes contained within them 228 , 614 (Figure 16.5).
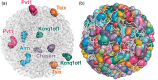
RNA promotes the formation of spatial compartments in the nucleus. (a) A 3D space filling nuclear structure model of selected lncRNAs. (b) A 3D space filling nuclear structure model of 543 lncRNAs that display at least 50-fold enrichment in the nucleus. (more...)
X-chromosome dosage compensation in Drosophila requires the formation of a phase-separated coacervate by the lncRNAs roX1 and roX2 interacting with the IDR of a specific partner protein (MSL2, ‘male sex lethal 2′). Moreover, replacing the IDR of the mammalian ortholog of MSL2 with that from Drosophila along with expression of roX2 is sufficient to nucleate ectopic dosage compensation in mammalian cells, showing that the roX–MSL2 IDR interaction is the primary determinant for compartmentalization of the X chromosome, and a likely exemplar of lncRNA-IDR interactions in general. 424
As further evidence that the eukaryotic nucleus (and indeed the eukaryotic cytoplasm) is finely organized, and that many more phase-separated domains remain to be discovered, a recent report has shown that two related RNA modification enzymes that normally reside (in one case) in the nucleolus and (in the other) in an unknown cytoplasmic domain proximal to mitochondria, both relocate upon nerve cell depolarization to different small unknown punctate nuclear domains. 703
The complexity is extraordinary and literature on this topic is burgeoning, but it is now clear that PSDs comprise a major and until recently unappreciated fine-scale and dynamic spatial regulation of subcellular and chromatin organization, “the active chromatin hub”, first proposed by Wouter de Laat and Frank Grosveld in 1993 based on the study of globin enhancers, 704 which “unifies the roles of active promoters and enhancers”. 520 It has also been proposed, with experimental support, that “ribonucleoprotein complexes can act as block copolymers to form RNA-scaffolding biomolecular condensates with optimal sizes and structures in cells”. 634
The ability of RNA to nucleate phase-separated domains adds a third dimension to its role in the origin of life. 705 While it has been widely accepted that RNA was likely the primordial informational and catalytic molecule of life, its advent would also have enabled the formation of a pre-cellular phase-separated privileged environment wherein organic reactions could be concentrated and evolve. Indeed, compartmentalized RNA catalysis has been demonstrated in membrane-free coacervate protocells. 706 , 707
The development of RNA-nucleated coacervates likely involved its interaction with positively charged (particularly arginine-rich) disordered proteins. 708 Intrinsically disordered proteins are encoded by the most ancient codons and appear to be the first polypeptides, likely to have functioned initially as chaperones, with catalysis transferred first from RNA to ribonucleoprotein complexes and then to proteins, 554 , 555 , 557 a process that may have been interactive. 709
The length of lncRNAs varies enormously although in many cases their true length and structure are unknown due to their cell-type specificity and low representation in RNA sequencing datasets. However, high depth sequencing has shown that most are multi-exonic, 710 and some are over 100 kb in length (post splicing), so-called macroRNAs, which have a mean length of 92 kb and are predominantly localized in the nucleus. 711
Why are lncRNAs so long? The likely answer is that they contain a set of modular domains for binding proteins and guiding them to target sequences in DNA or (other) RNAs 228 , 712–714 (Figure 16.6).

The modular domain structure and interactions of lncRNAs. (Reproduced from Mercer and Mattick.)
First, although rapidly evolving (under relaxed structure-function constrains and positive selection for adaptive radiation), lncRNAs exhibit common motifs and motif combinations across vertebrates, 715 and at least 18% of the human genome is conserved at the level of predicted RNA structure. 716 For example, it has been shown that conserved pseudoknots in lncRNA MEG3 are essential for stimulation of the p53 pathway. 717
Second, similar and potentially paralogous predicted RNA structures occur at many places throughout the genome. 718–720
Third, lncRNAs are enriched for repeat sequences, which have highly non-random distributions in them. 131 , 681 , 721 A notable feature of Xist, for example, is that its most highly conserved sequences are the repeat elements, whereas its unique sequences have evolved rapidly, 722 and many of its biological functions, including PRC2 binding, are mediated through its modular repeat elements. 371 , 379 , 380 , 387 , 397 , 723
Many of the lncRNAs referred to earlier have also been shown to be modular, with common features being rapid sequence evolution and structural divergence while retaining related functions, sometimes across large evolutionary distances, and the use of TE-derived sequences as protein-binding domains. 410 , 721 , 724 , 725
Indeed, TE-derived sequences and tandem repeats participate in many RNA-protein interactions, 34 , 675 , 726 , 727 which leads to the reasonable conclusion that repeat sequences act as RNA-, DNA-, and protein-binding domains that are the essential components of lncRNA function, 721 and that TEs are key building blocks of lncRNAs 397 , 728 (as well as fulfilling many other modular functions in gene control and gene expression; Chapter 10). Transposition is an efficient means of mobilizing functional cassettes 270 and allowing evolution to explore phenotypic space by modulation of the epigenetic control of developmental trajectories. As pointed out by Neil Brockdorf, “tandem repeat amplification has been exploited to allow orthodox RBPs [RNA binding proteins] to confer new functions for Xist-mediated chromosome inactivation … with potential generality of tandem repeat expansion in the evolution of functional long non-coding RNAs”. 397
Fourth, many lncRNAs bind chromatin-modifying proteins, transcription factors, nuclear matrix proteins and RNA-binding proteins, in those cases that are well studied, like Xist, roX and HOTAIR, to exert functional consequences. 309 , 311 , 342 , 371 , 727 It has also been reported that an mRNA can act as a scaffold to assemble adaptor protein assemblies to regulate intracellular transport. 729
Fifth, chemical probing has shown that lncRNAs, including Xist, physically have a modular structure, 371 , 374 , 730 , 731 and the chemical data matches that predicted by evolutionary conservation of secondary structure, validating both. 374
Finally, the extensive alternative splicing of lncRNAs strongly imputes a modular structure 710 , 732–735 and alternative splicing has, unsurprisingly, been shown to alter the function of lncRNAs. 735–738
If the complex ontogeny of a human requires a large number of guide RNAs then it is not surprising that many have similar protein-binding modules, with variation in repertoire and genomic target specificity, which may only require short stretches of nucleotide complementarity, given the high strength of RNA-RNA and RNA-DNA interactions. 739 These modules may also include enzymes that have adjunct roles in epigenetic transactions: for example, the developmentally regulated lncRNA H19 binds to and inhibits S-adenosylhomocysteine hydrolase, a feedback inhibitor of DNA methylyltransferases. 740 The alternative splicing of lncRNA exons (which itself must be epigenetically controlled 741 ) permits selection of specific protein-binding modules and target sites for feed-forward control of protein and regulatory RNA gene expression at many different loci and ultimately cell fate (hold, divide or differentiate) decisions at every stage of developmental ontogeny. There is no cogent model for such fine control by proteins alone.
The challenge now is to determine the repertoire of RNA structures using RNA folding programs, evolutionary conservation, physical and chemical analyses, and machine learning. 374 , 716 , 718–720 , 742–747 The challenge is to determine which RNA structures bind which proteins or which DNA or RNA targets, with a range of techniques becoming available to map RNA localization and interactions, 13 , 371 , 518 , 614 , 748–755 so that lncRNA biology can be parsed and understood, and thereby construct an expanded Rfam (‘RNA family’) database, 756 like the Pfam protein domain database 757 that has proved so useful in identifying protein function.
It is increasingly evident that lncRNAs, enhancers, topologically associated chromatin domains, transposon-derived sequences and other repeats, chromatin-remodeling and epigenetic information are merging into the same conceptual and mechanistic space.
Imagine the versatility and temporal precision that could be achieved if the locus specificity or local accessibility of proteins that control gene expression during development is guided by stage-specific modular RNAs. The strength of RNA is its potential to address targets through sequence-specific duplex or triplex base-pairing while at the same time recruiting and directing effector proteins to specific genomic locations.
We propose that regulatory RNAs, including the ‘repeat’ sequences within them, are the evolutionarily and developmentally flexible platforms expressed from the genome in an unfolding symphony to direct and execute the extraordinarily complex decisions required for the precise ontogeny of trillions of differentiated cells in a human, and similarly in other mammals, vertebrates, invertebrates and plants. That is, the epigenetic marks that control development, physiological adaptations and brain function are positioned and controlled by RNAs (among their many other functions in cell biology and gene regulation), and that the proportion of the genome devoted to specifying regulatory and architectural RNAs increases with developmental and cognitive complexity. 758 , 759
Small RNAs (miRNAs, sgRNAs, etc.) are simple sequence-specific guides for a single type of effector, such as RISC or CRISPR/Cas. LncRNAs not only have target sequence specificity but are also scaffolds for a range of proteins, notably chromatin-modifying complexes, with both targets and cargoes being four-dimensionally regulated by alternative splicing of lncRNA exons in a feed-forward cascade that directs the next cell fate decision during development. This is a highly efficient system that, like RNAi and CRISPR but in a far more sophisticated and modular manner, directs generic protein effectors to their sites of action.
The misleading historical perspective on the relationship between RNAs and proteins was best expressed by Ewa Grzybowska and colleagues, who concluded: “The current perception of RNA-protein interactions is strongly biased toward a protein-centric approach, in which proteins regulate the expression and activity of RNA, not the other way around.” 563
So-called because they have a domain shaped like a finger that is structured by a coordinated zinc ion.
Frequently mutated in pediatric kidney tumors and urinogenitary developmental disorders. 177
A subset of which can be specifically addressed by exact match to a transacting RNA, either a small RNA or an RNA sequence within a longer RNA.
Interestingly, the stability of genomic RNA-DNA hybrids in vivo is controlled by methylation of the RNA (see below). 197
There are other alternative and multi-stranded structures in eukaryotic genomes, including Z-DNA (binding domains for which occur in RNA editing enzymes, see Chapter 17), G-quadruplexes, I-motifs and cruciform structures, which, regrettably, despite the availability of specific antibodies, have not been mapped in genome-wide studies of genomic features and their dynamic relationship to cell type. 208–212 Many of these alternative DNA structures are formed by simple sequence repeats, which also abound in the genomes of plants and animals. 213
KZFPs (KRAB domain-containing zinc finger proteins) control the pleiotropic activation of TE-derived transcriptional cis-regulator sequences, some of which are primate-specific, during early embryogenesis, in part through histone H3K9me3-dependent heterochromatin formation and DNA methylation. 260 , 261 Primate-specific KZFPs also regulate gene expression in neurons. 262
Transcriptional gene silencing can be induced by siRNAs in the absence of DNA methylation, 286 indicating that small RNAs participate in other pathways that control chromatin state and architecture.
ANRIL also binds PRC1.
For many reasons, PRC2 has been the most intensively studied of all of the histone-modifying complexes with respect to the role of lncRNAs in epigenetic regulation of gene expression. 339
The presence of intrinsically disordered domains in most proteins involved in development has also been described as conferring promiscuity, a functional trait that allows flexible interactions in regulatory networks (see below).
LncRNAs also control the methylation of a number of non-histone proteins involved in cell signaling, gene expression and RNA processing. 365
Due to incomplete characterization of the enhancer RNA/transcription unit.
Evf2, which has a human homolog, also interacts with Sox2 to alter its target specificity, regulates transcription of the homeodomain transcription factors Dlx5 and Dlx6 as well as cohesin binding, and influences chromatin remodeling in the formation of GABA-dependent neuronal circuitry. 122 , 145 , 473 , 499
WDR5 is a a core subunit of the human MLL1-4 histone H3K4 methyltransferase complexes.
‘RNA regulates the formation, identity, and localization of phase-separated granules’. 542
p53 target gene association with nuclear speckles is driven by p53. 625
Japanese for ‘spotted pattern’.
PSDs may also serve to reduce noise in biological signal processing and control. 656
Interestingly, a similar enzyme in bacteriophage has been reported to add entire RNA chains to a host ribosomal protein to modulate the phage replication cycle. 691
Open Access: This content is Open Access under the Creative Commons license CC-BY-NC-ND.
Your browsing activity is empty.
Activity recording is turned off.
See more...